Weed identification and correct herbicide selection are essential for efficient weed management programs; however, selective herbicides may result in limited or no control without proper species identification. The primary mechanism for identifying weeds is through morphological and botanical features. However, weed identification in closely related species using physical characteristics may require botanical expertise, and the subtle differences between species can be complicated by further variations within subspecies.
A particular example is yellow foxtail and knotroot foxtail, which are problematic weed species in turfgrass that are often mistaken for one another due to their similar botanical characteristics (2, 5, 6, 7, 9). Accurate identification of these two weeds at the early growth stage is challenging, as key traits such as rhizomes are absent or appear late, long after identification is required for herbicide management (8). Although yellow and knotroot foxtail share similar morphological characteristics, they respond differently to several commonly used turfgrass herbicides, underscoring the need for accurate differentiation between the two species prior to herbicide application. The need for early and accurate differentiation is critical because yellow and knotroot foxtail can infest all major warm-season turfgrass species in the U.S., herbicide options are limited, and not all products are labeled or safe for use across all warm-season turfgrass species. To address this challenge, novel molecular technology has demonstrated great potential for rapid and reliable plant identification.
Figure 1. Knotroot foxtail (Setaria parviflora; left) with purple coloration above the crown and branching from the base. Yellow foxtail (Setaria pumila; right) at early stage. Photos by Mikerly Joseph

Nucleic acids are obvious options for identifying species across biological systems when physical identification and morphological identification is inconclusive. DNA barcoding is one molecular technique for identifying species by analyzing a short, standardized segment of DNA, similar to how a barcode identifies products (1). Different genes’ regions have been extensively used in plant DNA barcoding, and the most common are atpb-rbcL, matK, atpF-atpH, psbK-psbI, 18SF-26sR, trnH-psbA and nuclear ribosomal internal transcribed spacer (ITS) (4). However, there is no universal, one-size-fits-all approach, as sequence limitations or alignment issues in certain regions can produce inconclusive results (3).
Materials and methods
Research was conducted to evaluate the use of genetic markers for differentiating biotypes of yellow foxtail, knotroot foxtail, giant foxtail and green foxtail collected in Alabama. Biotypes of yellow and knotroot foxtail were collected in turfgrass, sidewalks, roadsides and pastures across Alabama, with 22 biotypes of yellow foxtail, 25 knotroot foxtail and seven classified as unknown. The USDA/ARS National Plant Germplasm System provided four other samples: yellow foxtail, knotroot foxtail, green foxtail and giant foxtail, which were our reference samples in the study.
Relevant sections of each plant, such as ligule, leaf size, leaf hair, seedhead size and roots system, were scrutinized and captured using Canon t5 camera with different macro lenses and Meiji EMZ dissecting microscope.
For genetic analysis, DNA extraction was performed at the Herbicide Resistance Diagnostics Lab at Auburn University, and extracted DNA from 48 biotypes was amplified with PCR targeting four specific barcoding regions — trnH-psbA, atpB-rbcL, matK and ITS — and sent for DNA sequencing. The DNA regions resulting from the sequencing were combined, and the Maximum Likelihood (ML) method alongside the General Time Reversible model were used to infer evolutionary history, utilizing 1,000 bootstrap replicates.

Figure 2. Setaria parviflora (left) presents with leaf hairs only near ligule, while Setaria pumila (right) presents longer leaf hairs along the entire leaf.
Results
Considering the morphological characteristics, yellow foxtail closely resembles knotroot foxtail, and standard measurements of culm lengths, blade widths, panicle and floret lengths often overlap between species, making identification a challenge. No single character taken on its own is sufficient for distinguishing the two species, as it takes evaluation of multiple characters at once. We found that yellow and knotroot foxtail have similar root systems at the early stage (Figure 1); however, knotroot foxtail is branched from the root base, while yellow foxtail is branched above the base. At the mature stage, yellow foxtail is annual with fibrous roots, while knotroot foxtail is perennial with rhizomes. The presence or absence of a hardened rhizome is perhaps the best characteristic for distinguishing the two species; however, the rhizome is sometimes not apparent. Therefore, if roots are not collected and inspected closely, along with other key characteristics, one may not arrive at a satisfactory identification. Knotroot foxtail stems are more geniculate, meaning they can bend in a slight zig-zag pattern, while yellow foxtail is sometimes bent only from the base. Regarding pubescence, knotroot foxtail has hairy ligules and long hairs near the leaf base (Figure 2). In contrast, yellow foxtail presents uniform leaf hairs along the entire leaf. Yellow foxtail seedheads are bigger and denser, with more hair than knotroot foxtail (Figure 3). However, the seedhead length and the height of the two species can overlap. The upper or fertile lemmas in knotroot foxtail are finely transversely rugose (cross-hatched patterned), compared with yellow foxtail, which has much more coarsely transversely rugose upper lemmas. In practice, plants of knotroot foxtail are generally smaller in many measurements when compared to individuals of yellow foxtail.
Figure 3. Seedhead of Setaria parviflora (left) and Setaria pumila (right) bigger and denser.

Considering the genetics analysis (Figure 4), by comparing the ITS region of the sample to the USDA check sample, yellow and knotroot foxtail share 98% of similarities, but yellow foxtail showed 96% and 92% similarities to giant foxtail and green foxtail, respectively. Knotroot foxtail showed 95% and 97% similarities to giant foxtail and green foxtail, respectively. We observed similar results in atpB-rbcL and matK regions, with higher similarities between yellow and knotroot foxtail than a single comparison of those to giant and green foxtail. No genetic variation was identified in the TrnH-psbA region. The ITS region discriminated yellow and knotroot foxtail with 16 SNPs detected. The atpB-rbcL region performed less with 18 SNPs, followed by matK with 7 SNPS. Additionally, in the atpB–rbcL region, we identified a consecutive 6-base-pair deletion at positions 160–166 bp in knotroot foxtail. In contrast, these base pairs are present in yellow foxtail, serving as a clear genetic marker to distinguish between the two species. To capture the evolutionary relationship between the species, ITS, atpB rbcL and matK were concatenated (2269–2289 bp) for Maximum Likelihood analysis. The result grouped the 48 samples into three main clusters: giant foxtail in one clade, green foxtail in another and a monophyletic yellow–knotroot clade. Within that clade, yellow and knotroot foxtail formed a specific sub group strongly supported by a bootstrap value of 99, cleanly separating them from green and giant foxtail in agreement with their closer genetic relationship. The barcode set (ITS, atpB rbcL, matK) provided robust, early stage resolution where morphology alone is ambiguous.
Conclusion
Yellow and knotroot foxtail are challenging to differentiate at an early stage. No single characteristic is sufficient for distinguishing the two species. Yellow and knotroot foxtail respond differently to selective herbicides, such as pinoxaden and sethoxydim, making accurate differentiation crucial prior to herbicide application in warm-season turfgrass. DNA barcoding provides an essential tool for species identification beyond morphological characteristics, conclusively proving that yellow foxtail and knotroot foxtail are two distinct genetic species. The application of the ITS, atpB-rbcL and matK DNA barcodes is highly relevant for accurately identifying foxtail species early in their development when morphological identification is inconclusive.
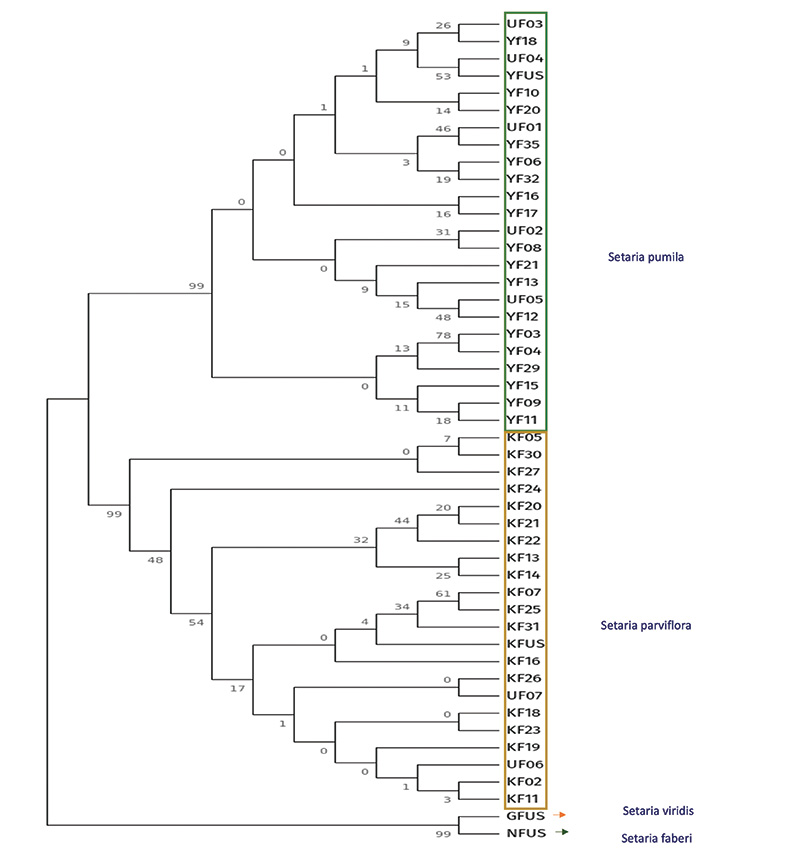
Figure 4. Phylogenetic tree developed using Maximum Likelihood method, Neighbor-Joining and visualized using Interactive Tree Of Life (iTOL). KF: Setaria parviflora. YF: Setaria pumila. UF: Unknown Setaria, GF: Setaria viridis. NF: Setaria faberi. YF, KF, UF followed by number are sample collected. YF, KF, GF, NF followed by US are sample check received from USDA- Germplasm Resources Information Network.
The research says
- Yellow and knotroot foxtail respond differently to selective herbicides, such as pinoxaden and sethoxydim, making accurate differentiation crucial prior to herbicide application in warm-season turfgrass.
- DNA barcoding provides an essential tool for species identification beyond morphological characteristics, conclusively proving that yellow foxtail and knotroot foxtail are two distinct genetic species.
- The application of the ITS, atpB-rbcL and matK DNA barcodes is highly relevant for accurately identifying foxtail species early in their development when morphological identification is inconclusive.
Literature cited
- Antil, S., J.S. Abraham, S. Sripoorna, S. Maurya, J. Dagar, S. Makhija, P. Bhagat, R. Gupta, U. Sood, R. Lal and R. Toteja. 2023. DNA barcoding, an effective tool for species identification: A review. Molecular Biology Reports 50(1):761-775 (https://doi.org/10.1007/s11033-022-08015-7).
- Bryson, C.T., and M.S. DeFelice. 2009. Weeds of the South. University of Georgia Press.
- CBOL Plant Working Group, P.M. Hollingsworth, L.L. Forrest, J.L. Spouge, M. Hajibabaei, S. Ratnasingham, M. van der Bank, M.W. Chase, R.S. Cowan and D.L. Erickson. 2009. A DNA barcode for land plants. Proceedings of the National Academy of Sciences 106(31):12794-12797 (https://doi.org/10.1073/pnas.0905845106).
- Chiang, T.-Y., B.A. Schaal and C.-I. Peng. 1998. Universal primers for amplification and sequencing a noncoding spacer between the atpB and rbcL genes of chloroplast DNA. Botanical Bulletin of Academia Sinica 39.
- Darmency, H., and J. Dekker. 2011. Setaria. Pages 275-291. In: C. Kole, ed. Wild Crop Relatives: Genomic and Breeding Resources. Springer.
- Dekker, J. 2003. The foxtail (Setaria) species-group. Weed Science 51(5):641-656 (https://doi.org/https://doi.org/10.1614/P2002-IR).
- Kellogg, E.A. 2017. Evolution of Setaria. In: A. Doust & X. Diao, eds. Genetics and genomics of Setaria (Vol. 19). Springer (https://doi.org/10.1007/978-3-319-45105-3).
- Joseph, M.M., B. Johnson, D.P. Russell, D.Y. Han and J.S. McElroy. 2024. Herbicide options for control of yellow and knotroot foxtail for possible use in turfgrass. Weed Technology 38(1):1-7 (https://doi.org/10.1017/wet.2024.45).
- Rominger, J.M. 1962. Taxonomy of Setaria (Gramineae) in North America. University of Illinois at Urbana-Champaign.
Mikerly Joseph (mikerlyjoseph@ufl.edu) is a graduate research assistant at the University of Florida, Gainesville; Jinesh Patel is an assistant research professor; Curtis Hansen is a herbarium curator, and J. Scott McElroy is a professor, all at Auburn University, Auburn, Ala.